Fossil Corkscrews after the Flood
Arphaxadian Paleontology
Chad Arment (2021)

From Barbour (1894), excavating daimonelices in Nebraska
Dr. Erwin Hinckley Barbour first examined the giant spiral-shaped fossils in northwestern Nebraska’s Miocene sediments in 1891. The ranchers and settlers of the region called them ‘fossil twisters,’ or ‘Devil’s corkscrews,’ inspiring Barbour to name them Daimonelix, sometimes spelled Daemonelix in his writings. (They are termed daimonelices in plural.) Barbour believed that these were fossilized plants with upright helical roots and roughly horizontal ‘rhizomes’ (though briefly considered that they might be freshwater sponges). Some scientists agreed with him, though a few thought they were simply concretions. Paleontologists E. D. Cope and Theodor Fuchs independently suggested that they could be rodent burrow casts, and this idea gained traction as Palaeocastor (a small burrowing beaver) remains were often discovered inside the fossil spirals. Even as late as the early 1940s, some were still arguing for a botanical origin (Lugn 1941), but Schultz’s (1942) review cemented the burrowing beaver theory as most probable.
These fossil burrows are trace fossils, also known as ichnofossils. As such, they have a taxonomic naming system that is separate from that which names fossilized organisms. Because burrows are sedimentary structures created by a living organism, they are also a type of lebensspuren (Martin and Bennett 1977). Barbour (1892) noted that a large number of these fossils covered an area of several square miles, all upright in position. Schultz (1942) pointed out that the abundance and distribution of Daimonelix suggested the rodents lived in colonies similar to modern prairie dog towns. Martin and Bennett (1977) noted that the highest concentrations of these fossils are in the Harrison Formation (Arikaree Group) of western Nebraska and eastern Wyoming.
The burrows are made up of two parts: a lengthy upright, spiral coil (which may be dextral or sinistral, according to Schultz (1942)), ending in a tubular burrow that often slopes up from the bottom of the spiral (possibly for effective drainage in heavy rains). Martin and Bennett (1977) noted that the helical burrows reach a depth of up to about 2.75 m (or 9 ft). Schultz (1942) noted that Palaeocastor remains are usually found in the lower chambers, where they likely formed nesting areas. The lower chambers can be as long as 4.5 m in length (Martin and Bennett 1977).
Martin and Bennett (1977) showed that the burrow walls provided an optimal micro-habitat for plant roots, allowing them to grow into a dense mat that helped maintain their stability, and even aided fossilization as, “Remarkable preservation of daimonelices . . . is shown to be due to rapid silification of plant roots invading the burrows.” An active burrow held root intrusion in check. When a burrow was abandoned, the root system packed the burrow full.
Calede (2013) noted that the subfamily Palaeocastorinae is comprised of several genera of fossorial (burrowing) or semi-fossorial beavers from the Oligocene and Miocene of North America. The genus Palaeocastor includes species adapted for scratch-digging and/or chisel-tooth digging (with ontogenetic changes likely influencing some shifts in behavior). Palaeocastor fossor is the species most commonly associated with daimonelices (Martin and Bennett 1977), though the larger P. magnus is also found with them—the two species did not, however, share ‘towns.’ Their paleoenvironment was “semi-arid, upland . . . sandy substrate,” and “never near evidence of ponded water” (Martin and Bennett 1977). They likely occupied open grasslands, grazing as do modern prairie dogs. Size-wise, P. fossor was about the size of a prairie dog.

Palaeocastor, from Scott (1962)
Why helical burrows?
Meyer (1999) provided a possible reason for these Palaeocastor species building corkscrew tunnels: “Helical burrows are more likely a response to palaeoclimates. The extra and/or unusual effort for the helical passage did result in corresponding increases in both burrow subsurface air volume and burrow wall surface area and/or in limiting air circulation. Those results would have provided greater thermal exchange that would have damped fluctuations in the burrow environment. Such environmental advantages through increased heat loss or gain would result in more consistent subsurface temperature and humidity for the helical design over a straight design when there were seasonal or extreme surface conditions.”
Doody et al. (2015) noted other possible reasons for long helical shafts before evaluating the helical burrows built by an Australian monitor lizard, Varanus panoptes. Those burrows reach about 3 m in depth. Eggs are laid in the nesting chamber at the bottom, and then most of the burrow is plugged up with dirt. As some pocket gophers build (smaller) helical burrows often associated with nesting, Doody suggested this could be a primary function of helix tunnels in general.
In a follow-up study, Doody et al. (2018) reiterated that no consensus had been made on the function of the corkscrew tunnels, but suggested that the extreme depth of the tunnels was more likely due to arid conditions rather than temperature. (A monitor lizard living in a more arid habitat dug deeper tunnels than a species living in a wetter climate.) As Palaeocastor likely lived in an arid grassland environment, deeper tunnels may have been beneficial to humidity control, even though those burrows were open, unlike the varanid burrows. (Of course, Martin and Bennett (1977) also noted humidity control as a possible factor, as modern fossorial rodent burrows can be saturated with moisture independent of burrow temperature or even soil moisture.)
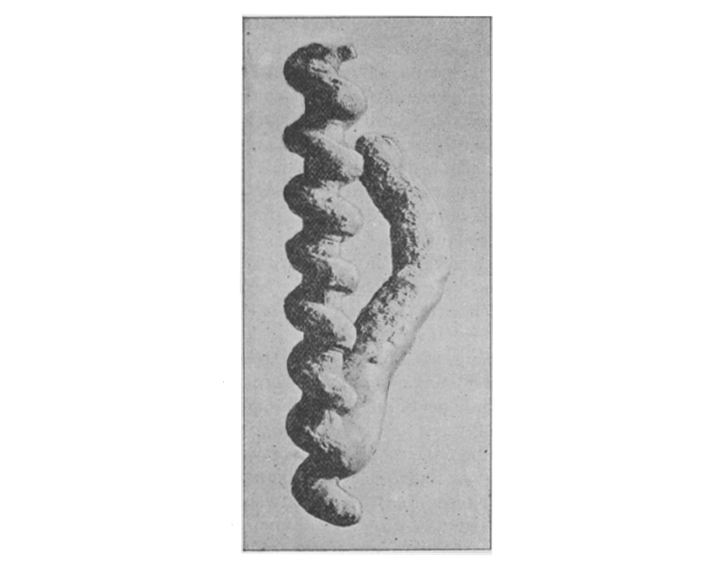
Barbour (1895)
When were these burrows formed?
I believe the evidence is overwhelmingly in favor of Palaeocastor burrows being trace fossils of the post-Flood era that creationists refer to as the Arphaxadian period (between Noah’s Flood and the Ice Age). The only alternatives to this would be a) the burrows were part of the pre-Flood topology and managed to survive the global flood intact, becoming fossilized in the process, or b) the burrows were formed during an intermittent stage within the global flood itself.
Could the burrows be pre-Flood relics? First, because they are lebensspuren, by definition they are essentially non-transportable (Frey 1975; Martin and Bennett 1977). Lebensspuren can be buried, but not moved without destroying the structures. In general, only a few trace fossils like coprolites and gastroliths can be transported. Body fossils, of course, are often found transported. The Harrison sediments are volcaniclastic, containing “substantial quantities of volcanic ash” (Martin and Bennett 1977). The NPS (2009) interpreted Harrison paleosols (the term for ancient soils) as “undifferentiated stream and eolian deposits.” Barbour (1892) characterized the fossilized sediments as “incompletely lithified sandstone,” noting the “gray matrix readily weathers away from the specimen.” This just does not fit into the narrative of a high-energy global flood. The paleosols suggest, rather, that the beavers were building burrows in a post-Flood sedimentary soil infused with volcanic ash from post-Flood volcanic activity.
This paleosol provided a solid foundation for post-Flood fossilization. Martin and Bennett (1977) noted that volcanic ash is a major source of amorphous silica. Ground water flowing through such an area leads to equilibrium with the silica. Dissolved and colloidally suspended silica attaches to organic matter and is readily absorbed by cellulose or lignin (e.g., the roots invading the burrow walls). The invading silica solution deposits silica throughout the burrow structure. “Thus a process of burrow diagenesis, involving a volcanic source area rich in amorphous silica and dependent upon the seasonal movement of ground waters in equilibrium with available silica, probably resulted in a very rapid partial lithification of beaver burrow walls, perhaps within a few years of the death of the plants occupying this microhabitat. Grass roots have been shown to decay fully in natural prairie soil within five years of the death of the plant. . . . Fossilization must have occurred before complete or even very extensive decay to account for the preservation of cellular detail. . . . It is now apparent that most of the preservation of burrow structures in the Harrison Formation is due to the infilling of the walls with roots which rapidly silicified” (Martin and Bennett 1977).
Second, the consistently upright position of the helical burrows in massive colonies, hundreds of known daimonelices at numerous fossil sites, Oligocene and Miocene, suggests they were not pre-Flood relicts. That would be an absurd reliance on statistical improbability.
Could they be rapidly-formed burrows during an intermittent stage within the Flood itself? While that might be possible with some fossil burrows (I’ll get to that below), it isn’t feasible for Palaeocastor burrows. Preservation of these burrows shows that plant roots infiltrated the burrow walls, and even took over entire burrows when they were abandoned. That obviously didn’t happen during the Flood; no plants grew during the Flood.
Very little attention has been given to these intriguing burrows from creationists. One anonymous (2001) creationist commentary on Meyer’s (1999) Palaeocastor paper noted: “Burrows of terrestrial vertebrates present an interesting problem for creationists. One would not expect animals to be constructing burrows during a flood, and their presence strongly indicates that the surface was exposed to the atmosphere during the time the burrows were being constructed.” As we’ll see, it is possible some burrows were formed during the Flood, but Palaeocastor burrows certainly were not. We have more creationist paleontologists now than we’ve ever had, and more in training, so hopefully the Arphaxadian period will receive more attention in the future.

Schultz (1942)
What about fossil burrows in Flood deposits?
We know that there are fossil burrows in Flood deposits, but there are significant differences from what is seen in Palaeocastor burrows. Here are a few that have been published:
1) A mid-Cretaceous sediment-filled burrow was found in Montana’s Blackleaf Formation with the remains of an adult and two juvenile hysilophodont (Oryctodromeus) dinosaurs (Varricchio et al. 2007). The burrow, sloping and sinuous in form, is more than 210 cm long, and 30-38 cm in diameter, until an enlarged cavity expansion of about 40-50 cm x 45 cm (where the bones are found). While the burrow is filled with sandstone, it cuts across three separate mudrock units. Varricchio et al. note that, “At least two episodes of filling would be required to account for the distinct sandstone units” that fill the burrow.
2) Two burrow casts were discovered in China’s Permian Naobaogou Formation, possibly formed by a dicynodont (fossil tetrapods found within the formation) (Liu and Li 2013). The largest was 1.6 m long, 23 cm wide, and 10 cm tall. The burrows are slightly curved in shape, excavated into mudstone and infilled with sandstone (a sandstone layer covering the mudstone). They are broken into pieces. The authors suggest that these burrows “indicate that the surface conditions are harsh or severely fluctuating and this area was semiarid or arid regions during that time.”
3) Tetrapod burrow casts were discovered in Antarctica’s Triassic Fremouw Formation and Lashly Formation (Sidor et al. 2008). All of the burrows were dug into “floodplain muds and subsequently filled by sand.” All nine burrows casts from the Allan Hills, Lashly Formation, were “collected from a single layer, which likely represents a pedogenic horizon.” (The authors interpret this as floodplain deposits that are not water-laden, which might confuse soil horizons through turbidity. From a creationist perspective, though, these burrows might suggest escape behavior within a very short time frame during the Flood.)
4) Fourteen cylindrical burrows were found in Utah’s Jurassic Escalante Member of the Entrada Sandstone (Loope 2006), with diameters up to 63 cm and lengths up to 3.05 m. They are roughly horizontal, and most were made in clusters. These burrows are interpreted as formed in sand dunes that would be too dry under normal conditions and “unsuitable for the semipermanent burrows needed for nesting and repeated sheltering.” When wet, such burrows could be easily created, but would soon collapse when they dried out. Loope suggests the burrows were “excavated in cohesive sand, presumably after precipitation events or wet seasons.” Exactly which organism created the burrows is unknown.
5) Five burrow casts were discovered in Poland’s Triassic deposits in the Holy Cross Mountains, possibly excavated by cynodont therapsids (Talanda et al. 2011). The burrows are in green and yellow mudstone and are filled with siltstone. The burrows are from 1 to 3.75 m in length and roughly descend from the horizontal at 18 to 36 degrees.
6) Helical burrow casts have been discovered in South Africa’s Permian Teekloof Formation, Beaufort Group, in the Karoo basin. Articulated skeletons of the small dicynodont Diictodon have been found in some, suggesting that it is the excavator (Smith 1987). Casts are usually broken up due to weathering on the surrounding mudrocks. Burrows are usually found in mudrocks and filled with sandstone or siltstone, though some are in sandstone and filled with mudrocks.
7) Short helical burrows (up to 71.4 cm deep) were found in Utah’s Jurassic Morrison formation, likely created by an unknown small mammal (Raisanen and Hasiotis 2018). They were superficially similar enough to Palaeocastor burrows to be named Daimonelix martini. (Within ichnotaxonomy, a trace fossils name is not necessarily linked to a specific organism, so two ichnofossils can share the same ichnogenus without being formed by two organisms of the same genus. There are other helical burrows given different names due to differing geometry.) These burrows (and another non-helical burrow form) are found primarily in carbonate-rich mudstone, preserved as carbonate fill (without linings or internal structures as seen in Palaeocastor burrows), which precipitates from the soil.
GRISDA’s anonymous creationist commentator (2001) noted, regarding a 2002 paper on Trirachodon burrows in the Triassic: “These burrows are in sediments that creationists generally regard to have been deposited during the Genesis Flood. Such features seem to indicate constrains on the nature of Flood activity in South Africa during the construction of these burrows. A possible explanation might be that the area was not under water at the time, but sediment was being intermittently washed into the area from some source area. Sufficient time passed between sedimentary episodes, such as flash floods, to permit burrow construction by these animals, which were similar in size to prairie dogs.”
This may be a feasible hypothesis for burrow excavation during the Flood—possibly a panic-induced burrowing response in certain organisms due to cataclysmic environmental conditions. In order to evaluate its potential, creation paleontologists would need to understand the nature of each of the sedimentary layers in question. Were muds and clays (in most cases above) present before the Flood, burrowed into, and then covered with additional sediment during the Flood, infilling the burrows with sands? That would make these pre-Flood burrows. Or were the muds and clays an early deposition of the Flood, burrowed into by surviving tetrapods, and covered with later sediments? That would make these peri-Flood burrows.
In any case, there are no serious similarities between these Flood-deposit burrow casts and Palaeocastor burrows. Creationists should interpret Palaeocastor burrows as part of the post-Flood landscape within the Arphaxadian period.
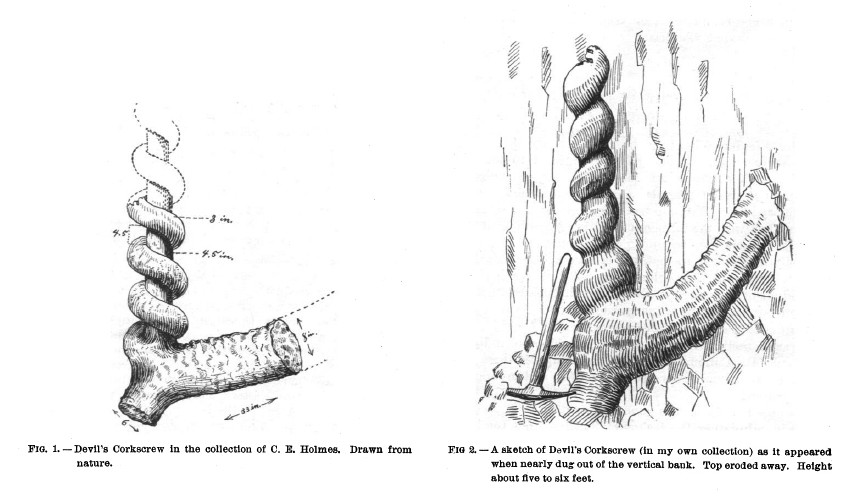
Corkscrew illustrations from Barbour (1892)
References
Anonymous. 2001. Annotations from the Literature. Origins [GRISDA] 52:28-47.
Bache, René. 1899. The Devil's corkscrews. The Strand Magazine 18: 593-595.
Barbour, E. H. 1894. Additional notes on the new fossil, Daimonelix. Its mode of occurrence, its gross and minute structure. University Studies (University of Nebraska) 2(1): 1-14.
Barbour, E. H. 1895. Is Daemonelix a burrow? A reply to Dr. Theodor Fuchs. American Naturalist 29(June): 517-527.
Barbour, E. H. 1897. History of the discovery and report of progress in the study of Daemonelix. University Studies (University of Nebraska) 2(2): 81-124.
Barbour, E. H. 1903. Present knowledge of the distribution of Daimonelix. Science N.S. 18(459): 504-505.
Barbour, I. H. [sic] 1892. Notice of new gigantic fossils. Science 19(472): 99-100.
Calede, J. J.-M. 2013. Skeletal morphology of Palaeocastor peninsulatus (Rodentia, Castoridae) from the Fort Logan Formation of Montana (early Arikareean): ontogenetic and paleoecological interpretation. Journal of Mammalian Evolution DOI 10.1007/s10914-013-9231-8.
Doody, J. S., et al. 2015. Deep nesting in a lizard, déjà vu devil’s corkscrews: first helical reptile burrow and deepest vertebrate nest. Biological Journal of the Linnean Society 116(1): 13-26.
Frey, R. W. 1975. The realm of ichnology, its strengths and limitations. in: Frey, R. W. (ed.) The Study of Trace Fossils. Springer: Berlin.
Liu, J., and L. Li. 2013. Large tetrapod burrows from the Permian Naobaogou Formation of the Daqingshan Area, Nei Mongol, China. Acta Geologica Sinica (English) 87(6): 1501-1507.
Loope, D. B. 2006. Burrows dug by large vertebrates into rain-moistened Middle Jurassic sand dunes. Journal of Geology 114: 753-762.
Lugn, A. L. 1941. The origin of Daemonelix. Journal of Geology 49(7): 673-696.
Mansfield, W. C. 1927. Some peculiar fossil forms from Maryland. Proceedings of the U.S. National Museum 71(2688): 1-9.
Martin, L. D., and D. K. Bennett. 1977. The burrows of the Miocene beaver Palaeocastor, western Nebraska, U.S.A. Palaeogeography, Palaeoclimatology, Palaeoecology 22(1977): 173-193.
Meyer, R. C. 1999. Helical burrows as a palaeoclimate response: Daimonelix by Palaeocastor. Palaeogeography, Palaeoclimatology, Palaeoecology 147(1999): 291-298.
National Park Service. 2009. Agate Fossil Beds National Monument: Geological Resources Inventory Report (2009/080).
Raisanen, D. C. W., and S. T. Hasiotis. 2018. New ichnotaxa of vertebrate burrows from the Salt Wash Member, Upper Jurassic Morrison Formation, south-eastern Utah (USA). Annales Societatis Geologorum Poloniae 88: 181-202.
Schultz, C. B. 1942. A review of the Daimonelix problem. University of Nebraska Studies [Studies in Science and Technology] 2(March): 5-30.
Scott, W. B. 1962. A History of Land Mammals in the Western Hemisphere. Revised edition. Hafner: New York.
Sidor, C. A., M. F. Miller, and J. L. Isbell. 2008. Tetrapod burrows from the Triassic of Antarctica. Journal of Vertebrate Paleontology 28(2): 277-284.
Smith, R. M. H. 1987. Helical burrow casts of therapsid origin from the Beaufort Group (Permian) of South Africa. Palaeogeography, Palaeoclimatology, Palaeoecology 60(1987): 155-170.
Talanda, M, S. Dzieciol, T. Sulej, and G. Niedzwiedzki. 2011. Vertebrate burrow system from the Upper Triassic of Poland. Palaios 26: 99-105.
Varricchio, D. J., A. J. Martin, and Y. Katsura. 2007. First trace and body fossil evidence of a burrowing, denning dinosaur. Proceedings of the Royal Society B 274: 1361-1368.
Daimonelices
Schultz (1942)
Daimonelices
Schultz (1942)
Daimonelix, Nebraska State Museum
Lugn (1941)
Daimonelices, Nebraska State Museum
Lugn (1941)
Daimonelices, Nebraska State Museum
Lugn (1941)
Excavation
Bache (1899)
Excavation
Bache (1899)
Excavation
Barbour (1894)
Excavation
Bache (1899)
Excavation
Bache (1899)
Original photo for Barbour (1894)
Agate Fossil Beds National Park / Wikipedia




